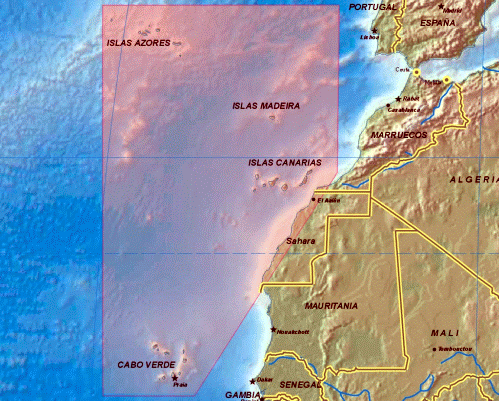
El sonoro término Macaronesia, de etimología griega (makáron =
felicidad, nesoi= islas) es utilizado por los estudiosos de la Naturaleza para
expresar un concepto fundamentalmente biogeográfico y botánico. Pero son los
botánicos los que aún no se ponen de acuerdo a la hora de limitar la extensión
territorial de esta región. Para muchos, quizás la mayoría, la Macaronesia
comprendería los archipiélagos noratlánticos de Azores, Madeira, Salvajes,
Canarias y Cabo Verde, además de una amplia franja costera africana situada
frente a dichas islas, que va desde Marruecos hasta Senegal. Esta extensa unidad
biogeográfica, esencialmente botánica, se encuentra entre las coordenadas: 39º
45’N, 31º 17’W de la isla más septentrional que es Corvo (Azores), y 14º
49’N, 13º 20ºW de Brava (Cabo Verde), la más meridional. Por otro lado,
Flores (Azores) es la más alejada del continente (a 1.900 km de Europa) y
Fuerteventura (Canarias) la más próxima (a 96 km de Africa). Como es lógico,
estas dispares situaciones geográficas propician una variabilidad climática
sensible, lo que ,a su vez, condiciona el poblamiento biológico. Y si a esto añadimos
las notorias diferencias en cuanto a distancias al continente entre unos archipiélagos
y otros, se hace evidente la necesidad de tener en cuenta todos esos factores a
la hora de hacer estudios y valoraciones biogeográficas y ecológicas.
En este contexto trataremos de dar una visión, lo más sucinta posible,
sobre el origen, situación geográfica, clima, flora y fauna de cada uno de los
archipiélagos y del enclave continental. De igual modo trataremos, de manera
general, y particular en su caso, de una serie de aspectos paleontológicos y
paleoecológicos que consideramos de interés para una mejor comprensión del
origen, poblamiento biológico y evolución de esta singular región del Atlántico
nororiental.
El
origen
 Los archipiélagos macaronésicos tienen en común su origen volcánico.
Todas las islas se pueden considerar como oceánicas, o lo que es lo mismo, han
emergido del mar tras sucesivas erupciones submarinas de magmas fundamentalmente
básicos (basalto), a través de fracturas y zonas de debilidad de la corteza
oceánica. Pero hay algunas islas, como Lanzarote y Fuerteventura, que al estar
más próximas al continente, se asientan sobre corteza de transición,
continental-oceánica, y por lo tanto, el magma en su ascenso puede arrastrar a
la superficie fragmentos de rocas sedimentarias del borde continental africano.
Los archipiélagos macaronésicos tienen en común su origen volcánico.
Todas las islas se pueden considerar como oceánicas, o lo que es lo mismo, han
emergido del mar tras sucesivas erupciones submarinas de magmas fundamentalmente
básicos (basalto), a través de fracturas y zonas de debilidad de la corteza
oceánica. Pero hay algunas islas, como Lanzarote y Fuerteventura, que al estar
más próximas al continente, se asientan sobre corteza de transición,
continental-oceánica, y por lo tanto, el magma en su ascenso puede arrastrar a
la superficie fragmentos de rocas sedimentarias del borde continental africano.
En cualquier caso, todos estos archipiélagos son una consecuencia de la
geodinámica del Océano Atlántico que -a través de las fracturas y fallas de
transformación y, fundamentalmente, de la cordillera dorsal- no ha dejado de
emitir magma desde su apertura hace más de 180 millones de años. Las
investigaciones oceanográficas de los fondos atlánticos en las últimas décadas
han revelado que su edad aumenta a medida que se alejan (1 ó 2 cm al año) de
la dorsal, en ambas direcciones. Y así resulta que en Azores hay dos islas
(Flores y Corvo) –las más jóvenes- que se están alejando del resto en
dirección a América, ya que se encuentran separadas por la dorsal atlántica.
De igual modo, los ejes estructurales principales de la geotectónica atlántica
inducen las direcciones y alineaciones insulares, en las que se observa un
predominio de las NE-SW y NNW-SSE, como lo demuestran los bancos submarinos
entre Madeira y la Península Ibérica, o la alineación Fuerteventura -
Lanzarote - isletas - Banco de Concepción.
Por
otra parte, a excepción de Azores, los demás archipiélagos macaronésicos se
encuentran en el interior de la placa tectónica africana, lo que les confiere
un menor riesgo sísmico y una más atenuada actividad volcánica. Sin embargo,
las islas Azores -situadas en el borde de la placa, a ambos lados de la dorsal,
con una clara alineación en la dirección estructural NNW-SSE, y próximas a la
zona de fractura del Atlántico oriental- presentan una lógica mayor actividad
volcánica y sísmica. En los últimos años han ocurrido allí, principalmente
en las islas centrales, seismos de intensidad destructiva que provocaron víctimas
mortales.
Las islas volcánicas oceánicas comienzan a poblarse desde el momento en
que emergen y se enfrían los materiales magmáticos que las han hecho nacer.
Existen tres agentes fundamentales que propician la dispersión genética y el
poblamiento insular: los vientos
(anemocoria) capaces de transportar a grandes distancias propágulos de
semillas, pequeños insectos y arácnidos, etc.; las
corrientes marinas (hidrocoria), que propician el transporte pasivo -por
flotación- de semillas, animales en balsas de poblamiento, etc., y favorecen el
transporte activo de otros animales poco nadadores (algunos mamíferos y
reptiles), y en tercer lugar las aves
(ornitocoria), que pueden transportar en su aparato digestivo, en las patas y
plumaje, toda una serie de frutos, semillas y pequeños insectos. No podemos
olvidar a los mamíferos voladores (murciélagos) que pueden llevar parásitos
en su pelaje, o a los insectos voladores que también pueden ser agentes de
transporte para otros artrópodos más pequeños (ácaros). Finalmente, debemos
considerar al hombre como un agente de dispersión biológica importante
(antropocoria), muchas veces nefasta.
Como es lógico, la distancia al continente o a otras islas próximas,
juega un papel importante en el poblamiento, lo mismo que la edad, situación
geográfica, clima, superficie y altura de las islas. Es, por lo tanto, la
combinación de todos estos factores lo que se debe valorar en cualquier
investigación sobre la biogeográfia y biodiversidad insulares. Por todo ello,
no nos debe extrañar que las islas Canarias, las más favorecidas en estos
condicionantes con respecto a los otros archipiélagos macaronésicos (mayor
extensión, menor distancia al continente, clima subtropical, mayor altura),
presenten las más altas tasas de biodiversidad. Recientes investigaciones
parecen apuntar a una colonización de “ida y vuelta” por parte de algunas
especies vegetales que llegaron a estas islas procedentes de África,
evolucionaron en ellas y ahora han regresado al continente ya diferenciadas. (Aeonium
spp.).
Pero
nos estaríamos olvidando de un factor fundamental en el poblamiento de la
Macaronesia, si no mencionamos el papel que han jugado los actuales bancos
submarinos que rodean a los archipiélagos, algunos –incluso- en sus aguas
interiores, los cuales han actuado como “puentes” de dispersión y
colonización, en las épocas en que esos bancos, como el de Ormonde,
Gettysburg, Ampere, Seine, Dacia, Concepción y otros, fueron islas, algunas de
superficie ya considerable (200-300 km2).
No
insistiremos demasiado en el poblamiento marino de las islas, debido a la mayor
capacidad y posibilidades de dispersión que tienen las especies a través de
las corrientes, sobre todo en su etapa larvaria, y tratarse el mar del medio
homogéneo y muy extenso que rodea a las islas. En cualquier caso, cabe decir
que el sistema de circulación de las corrientes y las condiciones oceanográficas
han variado con el tiempo, permitiendo el avance o retroceso de fauna y flora
desde zonas frías hacia los trópicos, o viceversa, como es el caso de la
presencia de fauna tropical en Azores durante el Cuaternario. También es
resaltable la gran capacidad de dispersión genética que tienen algunas
especies marinas para desplazarse a largas distancias en estado larvario, a través
de las corrientes. A título de ejemplo cabe decir que hemos detectado más de
120 especies de moluscos gasterópodos anfiatlánticos (que viven en las dos
orillas del océano), y lo mismo sucede con muchos peces, crustáceos,
equinodermos, algas, y otros grupos de fauna y flora marinas.
Finalmente,
nos resta señalar que hasta el siglo XV el único archipiélago macaronésico
habitado era Canarias, que fue colonizado en la antigüedad (primer milenio
A.C.) por pueblos norteafricanos. La llegada del hombre a cualquier isla
deshabitada siempre representa un trauma para la flora y fauna locales, no sólo
por la introducción del cortejo de especies domésticas que le acompañan
(cabras, perros, gatos, cerdos, etc.) y de vegetales para cultivo, sino por la
desforestación, urbanización, contaminación, caza incontrolada y, en
definitiva, destrucción de los ecosistemas naturales, efecto del que no se han
librado los archipiélagos macaronésicos.
Por
una serie de circunstancias u oportunidades ecológicas y procesos de genética
de poblaciones, aún no del todo desvelados, se ha constatado que en las islas
la evolución se dispara. En los ecosistemas insulares, sobre todo en islas oceánicas,
los fenómenos evolutivos de especiación o radiación adaptativa actúan con
mayor rapidez que en sus correspondientes continentales. Pero, al mismo tiempo,
esos endemismos se vuelven más vulnerables porque han perdido sus mecanismos de
defensa contra los competidores del continente. Una posible explicación a esto
la tenemos en el “efecto de fundación”, según el cual, un pequeño grupo
fundador, o pionero, de individuos pertenecientes a una especie recién llegada
a la isla, es portador de sólo una fracción de la información genética total
de la población originaria, de la cual se ha separado. Por lo tanto, los propágulos
de esta especie al estar privados de parte de su potencial evolutivo, tienen
necesidad de adaptarse lo más rápidamente posible a las nuevas presiones de
selección, muy diferentes de las que sufrió la población madre en el
continente.
Parece
que una de las salidas que encuentran esas especies en el medio insular es el
cambio de tamaño, observándose en muchos casos la tendencia tanto al
gigantismo como al nanismo, de tal manera que los pequeños herbívoros
(roedores, lagomorfos, tortugas terrestres) tienden a aumentar su tamaño
corporal, mientras que los predadores carnívoros (mamíferos y reptiles) lo
hacen a la inversa. Como es de suponer, estos cambios están relacionados con la
disponibilidad de recursos energéticos (alimento). En todo caso, la
vulnerabilidad es mayor en las especies de gran tamaño. En el registro fósil
de Canarias hay buenos ejemplos de ello, como son las tortugas gigantes fósiles
(Geochelone spp.) de Tenerife, Gran
Canaria, Fuerteventura y Lanzarote, el lagarto gigante (Gallotia
goliath) de Tenerife y La Gomera, y la rata gigante (Canariomys
spp.) de Tenerife y Gran Canaria. En suma, parece que ésta es una norma que
se repite en las islas a lo largo del tiempo. Basta comparar los dos archipiélagos
volcánicos de Galápagos y Canarias para darnos cuenta de que, en el primero
-mucho más jóven- está sucediendo ahora lo que ocurrió en Canarias hace
millones de años (grandes tortugas terrestres, iguanas-lagartos).
En
estos archipiélagos pequeños, verdaderos laboratorios de la evolución, la
presencia del hombre ha supuesto un golpe duro a sus ecosistemas y
biodiversidad. Su limitada superficie insular acentúa la fragilidad de los
mismos y su indefensión ante la arribada del gran “superdepredador”, hecho
que algunos comparan con el “Blitz-krieg” refiriéndose a la extinción del
70% de los grandes mamíferos americanos (mamuts, équidos, megaterios) que
evolucionaron en ese continente -el cual se comportó como una isla gigantesca-
hasta el fin de la última glaciación, hace 11.000 años, época en que grupos
de cazadores asiáticos cruzaron el estrecho de Béring. Igual sucedió con los
marsupiales de Tasmania, las moas de Nueva Zelanda, las aves elefante de
Madagascar, el dodó de Mauricio y las tortugas gigantes de las islas Mascareñas,
entre otros. En la Macaronesia la intervención humana ya se ha hecho notar con
la desaparición de la foca monje (Monachus monachus) y del milano (Milvus milvus) en Canarias, los grandes escíncidos de Cabo Verde (Macroscincus
coctei) o los dragos (Dracaena draco)
de Porto Santo (Madeira).
Paleoecología
 Creemos
interesante hacer unas consideraciones paleoecológicas, que puedan ayudar a
comprender mejor los procesos ecológicos actuales. La paleoecología estudia
las condiciones ecológicas que existían en el pasado, en base a datos
indirectos proporcionados por los caracteres de adaptación al medio en que
vivieron que presentan los fósiles. Para ello también hay que recurrir a la
naturaleza inorgánica, ya que la geomorfología, petrología, sedimentología y
otras ciencias, proporcionan valiosos datos sobre las condiciones
medioambientales de épocas pasadas. En todo caso, son los fósiles vegetales
los que mejor señalan los paleoclimas, pues al poseer las plantas menor
movilidad que los animales, acusan más los cambios climáticos y lo reflejan en
su estructura interna y externa. También cabe añadir que los ecosistemas
actuales representan sólo una parte de los que han existido a lo largo de todo
el tiempo transcurrido desde la aparición de la vida sobre nuestro planeta. Por
eso deberíamos hablar de perspectiva
ecológica, ya que al estudiar antiguas comunidades de plantas y animales, y
sus cambios a través del tiempo, es posible comprender más plenamente la
historia del desarrollo, estructura y función de los ecosistemas modernos.
Creemos
interesante hacer unas consideraciones paleoecológicas, que puedan ayudar a
comprender mejor los procesos ecológicos actuales. La paleoecología estudia
las condiciones ecológicas que existían en el pasado, en base a datos
indirectos proporcionados por los caracteres de adaptación al medio en que
vivieron que presentan los fósiles. Para ello también hay que recurrir a la
naturaleza inorgánica, ya que la geomorfología, petrología, sedimentología y
otras ciencias, proporcionan valiosos datos sobre las condiciones
medioambientales de épocas pasadas. En todo caso, son los fósiles vegetales
los que mejor señalan los paleoclimas, pues al poseer las plantas menor
movilidad que los animales, acusan más los cambios climáticos y lo reflejan en
su estructura interna y externa. También cabe añadir que los ecosistemas
actuales representan sólo una parte de los que han existido a lo largo de todo
el tiempo transcurrido desde la aparición de la vida sobre nuestro planeta. Por
eso deberíamos hablar de perspectiva
ecológica, ya que al estudiar antiguas comunidades de plantas y animales, y
sus cambios a través del tiempo, es posible comprender más plenamente la
historia del desarrollo, estructura y función de los ecosistemas modernos.
Cuando hablamos de paleoclimas debemos tener bien presente que durante épocas
húmedas del pasado, las islas volcánicas como éstas, con frecuencia se vieron
afectadas por intensas borrascas y grandes precipitaciones que produjeron una
fuerte erosión -muchas veces súbita- por aluviones que dieron lugar a
profundos barrancos, como los de Madeira, Santo Antao, Gran Canaria o La Gomera,
y a depósitos sedimentarios de notable potencia, como los del Valle de Güímar,
Tenerife. Sin embargo, durante las épocas de aridez la erosión es paulatina,
produciéndose una pérdida de masa vegetal y de suelo, emigraciones
altitudinales de la vegetación, si la orografía de la isla lo permitía y,
finalmente, extinciones.
Aunque ya hemos mencionado algunos acontecimientos naturales de
consecuencias catastróficas para la vida, conviene que insistamos en los
efectos que pudieron tener en los ecosistemas insulares. Si comenzamos con los
de origen geológico, los grandes deslizamientos de los valles de La Orotava
(hace 170.000 años) y Güímar en Tenerife, y el del Golfo en El Hierro, son
buenos ejemplos de cómo súbitamente puede desaparecer bajo el mar una gran
porción de la isla y alterarse drásticamente el equilibrio ecológico, en esa
isla y las vecinas, pues los tsunamis que originan estos deslizamientos son de
dimensiones colosales, con olas de más de 100 m de altura. Es muy posible que
muchas formaciones litorales de estas islas a las que no se ha conseguido dar
explicación, tengan su origen ahí. Pero estos fenómenos catastróficos no son
privativos de Canarias, pues sus resultados también se pueden observar en otros
archipiélagos macaronésicos como Cabo Verde (Fogo), y de otros lugares del
mundo (Tristán da Cunha, Hawaii, Juan Fernández, etc.).
Otros acontecimientos
catastróficos que han alterado la paleoecología insular son las erupciones
violentas y explosivas, de carácter ácido, con emisión de nubes ardientes y
coladas piroclásticas, que sepultan y aniquilan toda forma de vida que
encuentran a su paso, como les ocurrió a las grandes tortugas terrestres de
Tenerife (Testudo burchardi).
Las erupciones freatomagmáticas, con grandes explosiones, al entrar el
agua del mar en contacto con la cámara magmática, y la formación, en algunos
casos, de amplias calderas, alteran sensiblemente los ecosistemas locales. Tal
es el caso de la Caldera de Pedra de Lume en la isla de Sal (Cabo Verde), que
contenía en su interior- en épocas pasadas- una laguna de agua dulce como las
actuales de Azores, y ahora, al desecarse, es una salina.
Grandes incendios forestales o el impacto de asteroides en regiones
cercanas a las islas, como puede ser el de Richat en Mauritania, son
acontecimientos catastróficos que, con toda probabilidad, también alteraron la
naturaleza macaronésica en el pasado.
La gran mortandad de pardelas (Puffinus holei) ocurrida hace unos 30.000 años en Fuerteventura, así como otros yacimientos de difícil interpretación, quizás tuvieron su explicación en estos fenómenos.
El archipiélago de Azores, situado en medio del Atlántico, a unos 1600
km al oeste de Portugal y a 2700 de la costa oriental norteamericana, representa
a la Macaronesia verde y húmeda. Su disposición alineada con la dirección
NNW-SSE está en consonancia con una de las directrices estructurales de la tectónica
atlántica. Las nueve islas que lo componen se disponen en tres grupos: el
suroriental, constituido por San Miguel y Santa María, el central por Terceira,
San Jorge, Graciosa, Pico y Faial y el noroccidental por Flores y Corvo. Estas
islas le restan al océano una extensión de 2350 km2 , de los que
760 corresponden a la mayor, San Miguel, y 17 a Corvo, la más pequeña y
septentrional.
La proximidad del archipiélago de Azores al borde norte de la placa tectónica
africana y a la zona de fracturas del Atlántico norte, así como su situación
con respecto a la cordillera dorsal atlántica -que lo atraviesa y hace que
Flores y Corvo se estén separando del resto- es el motivo de que estas islas
registren una notable actividad volcánica y sísmica. Fruto de este volcanismo
activo son las fumarolas y solfataras (caldeiras) con emisión de gases
sulfurosos a gran temperatura, que provocan la ebullición del agua y lodo
acumulados en las depresiones de algunas zonas. Este volcanismo residual es
aprovechado, aunque de manera incipiente, en algunas pequeñas plantas de energía
geotérmica. De origen netamente oceánico, podemos asegurar que las Azores son,
en su conjunto, las islas más jóvenes de la Macaronesia, observándose que la
edad disminuye a medida que se acercan a la dorsal atlántica.
En Santa María, la más antigua, se encuentran depósitos fosilíferos
marinos del Mioceno superior, así como niveles cuaternarios (playas levantadas)
que contienen fauna tropical actualmente desaparecida de aquellas latitudes.
El clima de Azores es oceánico, templado y húmedo, influenciado por el
anticiclón que lleva su nombre. Sin embargo, es muy variable, destacando su
elevada pluviometría, que da origen a lagunas de considerable extensión en el
interior de calderas o depresiones de diferentes islas. Es muy famosa la de Sete
Cidades, en San Miguel, que con sus 5 km de diámetro y sus dos partes bien
diferenciadas (laguna verde y laguna azul), proporciona un espectáculo paisajístico
de extraordinaria belleza.
La temperatura del archipiélago está influenciada por la corriente cálida
del Golfo, que contribuye a suavizar los inviernos (14º C de media) y los
veranos (22º C).
Estos factores climáticos propician la existencia de una flora
esplendorosa de bosques y pastizales, pero lamentablemente la vegetación
natural autóctona ha sido sustituida en gran parte por especies introducidas.
Por tal motivo, de aquellos extensos bosques de laurisilva que en su día
existieron, no quedan sino pequeños relictos. En su lugar se han repoblado
grandes extensiones con una conífera de origen japonés (Cryptomeria
japonica).
En general, la flora endémica de Azores es más afín a la europea,
aunque presenta algunos endemismos que pertenecen a géneros bien representados
en la Macaronesia, como Pericallis,
Tolpis, Aichryson, Cedronella, Picconia, etc.
Una consecuencia lógica de su alejamiento del continente es la pobreza
de la fauna de este archipiélago, y además, la juventud del mismo hace que la
especiación insular no haya operado a lo largo de mucho tiempo, de tal manera
que los endemismos son escasos. No obstante, cabe destacar la presencia de razas
ornitológicas propias de estas islas, como el ratonero Buteo buteo rothschildi, la paloma torcaz Columba columba azorica y el mirlo Turdus merula azoriensis, o razas insulares como las del reyezuelo Regulus
regulus inermis, presente en Pico, Terceira, Faial, San Jorge y Flores, R.
regulus azoricus en San Miguel y R.
regulus santaemariae en Santa María.
Del resto de la fauna azoreana podemos destacar que los
diferentes grupos de invertebrados: insectos, arácnidos, moluscos, etc.,
muestran una mayor afinidad con la fauna europea que la que tienen los restantes
archipiélagos macaronésicos. Con la fauna y flora marina ocurre algo parecido,
a pesar de que Azores se ve bastante influenciado por la corriente de El Golfo.
El archipiélago de Madeira, compuesto por las islas de Madeira, Porto
Santo, Las Desertas y algunos islotes, se encuentra a unos 700 km del continente
africano y a 950 de las costas europeas, o lo que es lo mismo, en pleno dominio
oceánico. Este conjunto insular, de unos 800 km2, forma parte de un
extenso accidente estructural oceánico que va desde Cabo Verde hasta las costas
portuguesas, discurriendo más o menos paralelamente a la dorsal atlántica.
De indudable origen volcánico-oceánico como el resto de los archipiélagos
macaronésicos, Madeira, y especialmente Porto Santo, muestran formaciones
sedimentarias fosilíferas que han permitido su datación entre 10 y 15 millones
de años. En todo caso, se ha considerado que la causante del origen de estas
islas ha sido la presencia de una “pluma caliente” de larga duración,
procedente del manto subyacente al archipiélago.
Las edades más antiguas (18 m.a.) se encuentran en los niveles más
elevados del volcanismo submarino del complejo basal de Porto Santo, en donde
aparecen formaciones recifales intercaladas. Más joven es Madeira, en la que la
parte superior del complejo volcánico basal, que también presenta una
intercalación recifal del Mioceno, fue datada en 5.2 millones de años.
De las Desertas, más modernas aún (2-3 millones de años), destacaremos
la clara plataforma de abrasión que nos muestra Chao en la superficie (de ahí
su nombre) y que es el resultado de la intensa labor erosiva del mar en épocas
cálidas de finales del Cenozoico, en las que el nivel subió por encima de los
70 m.
Los vientos alisios del NE, junto a la corriente de Canarias, marcan el
clima del archipiélago de Madeira que, como ocurre en las demás islas macaronésicas,
es variable o se encuentra modificado por la orografía y orientación de cada
una de ellas. Así, el clima de Madeira, en líneas generales, es diferente al
de Porto Santo o al de las Desertas, ya que su altura (1860 m) y abundante
vegetación propician que las precipitaciones sean allí mucho más copiosas que
en las otras islas, llegando a sobrepasar, en las zonas más altas y húmedas,
los 2.800 mm. Con respecto a las temperaturas, señalar que son bastante
similares a las de Canarias, con escasas diferencias de 1 ó 2 ºC inferiores en
las medias, debido fundamentalmente a la superior latitud de Madeira.
Al hablar de la flora de este archipiélago debemos comenzar por el grave
deterioro que ha sufrido la vegetación desde el momento (siglo XV) en que el
hombre puso el pie en ellas. Basta decir que era tal la densidad forestal en
Madeira (llegaba hasta el mar) que hubo que prenderle fuego par poder penetrar
en la isla. De esta manera Madeira estuvo ardiendo, se dice, durante 6 años.
Ante este panorama, agravado tras la introducción de animales herbívoros, como
cabras, conejos, burros, vacas, etc., no es difícil imaginar el estado actual
de la flora autóctona de estas islas. En Madeira los restos de la vegetación
original se encuentran ahora refugiados en las zonas más inaccesibles y
escarpadas de las cumbres y valles (laurisilva), así como en los acantilados
costeros y montañas del piso basal: tabaibas (Euphorbia
piscatoria), dragos (Dracaena draco)
y sabinas (Juniperus phoenicea).
Destacan también los verodes o pasteles de risco (Aeonium spp.), abundantes en las paredes y taludes de la zona norte
de la islas.
En
Porto Santo es la vegetación xerófila la mejor representada, de acuerdo con
las condiciones geomorfológicas y climáticas de esta isla. Euphorbia
piscatoria encuentra en ella un hábitat más apropiado que en Madeira.
Cuentan
las crónicas y escritos antiguos que los dragos fueron abundantes en Porto
Santo, y hoy, lamentablemente, han desaparecido de la vegetación natural de sus
montañas.
En
las Desertas, como cabría esperar, la vegetación es escasa, conformada
fundamentalmente por plantas herbáceas y líquenes, y está muy deteriorada por
la presencia de cabras que, según cuentan, fueron traídas de Canarias después
de su conquista. Es muy posible que ésta sea la antigua cabra canaria prehispánica,
puesto que está constatado que un buen contingente de pastores guanches fue
trasladado a Madeira junto a numerosos rebaños caprinos.
La
fauna madeirense, al igual que en los demás archipiélagos, está en función
de algunos parámetros variables, como pueden ser la superficie de la isla, su
altura y, sobre todo, la distancia al continente. En este sentido podríamos
decir que Madeira se encuentra en una situación intermedia entre Canarias y
Azores, de ahí que sea pobre en algunos grupos, como la herpetofauna, ya que
los reptiles están entre los que mayores dificultades encuentran a la hora de
cruzar amplios brazos de mar. De tal manera que nos encontramos con un sólo
lagarto (Podarcis dugesii), que también vive en Salvajes y Azores, un gecónido
(Tarentola mauritanica) y algunos escíncidos
(Chalcides).
Pero
son las aves terrestres el grupo más representativo y abundante de vertebrados,
con más de 40 especies nidificantes, entre las que destacan especies endémicas
como la paloma de madeira (Columba trocaz),
o el petrel de Zino (Pterodroma madeira)
y subespecies insulares, como el reyezuelo (Regulus
ingnicapillus madeirensis), el pinzón (Fringilla
coelebs maderensis) o el ratonero (Buteo
buteo harterti). También están allí presentes endemismos macaronésicos
como el canario (Serinus canarius), el
vencejo (Apus unicolor) y el bisbita
caminero (Anthus bertheloti madeirensis).
Entre
los mamíferos autóctonos señalaremos los murciélagos, destacando el
endemismo macaronésico Pipistrellus
maderensis. No obstante, han sido los mamíferos introducidos por el hombre
los mayores devastadores de la fauna y flora de este archipiélago. Las ratas,
ratones, conejos, cerdos, cabras, gatos, etc., han producido un daño tremendo
en la biota insular, llevando al borde de la extinción a algunas especies autóctonas.
Las
islas Salvajes conforman un pequeño archipiélago constituido por tres isletas
y algunos roques. La mayor, Salvaje Grande, tiene una superficie de 4 km2,
Salvaje Pequeña o Gran Pitón, 0’5 y la Salvajita o Ilheu de Fora, 0’2 km2.
Estas pequeñas islas, situadas entre Madeira y Canarias, distan 160 km de
Tenerife y 250 de Madeira. Su mayor proximidad a Canarias hace que existan
grandes afinidades en su génesis, presentando un Complejo Basal intrusivo y
extrusivo datado entre 24 y 27 millones de años.
Las
Salvajes han estado sometidas a una
intensa erosión marina, que se manifiesta en Salvaje Grande por medio de una
plataforma de abrasión situada a unos 100 m de altura, sobre la cual se
depositaron sedimentos marinos fosilíferos. Estos depósitos sedimentarios a su
vez, fueron recubiertos posteriormente por coladas volcánicas producto de una
intensa actividad hace 11.5 millones de años. Luego de un prolongado período
de calma, ocurrió hace 4 m.a. una reactivación del volcanismo. Estas dos fases
magmáticas han tenido sus equivalentes, más o menos sincrónicas, en el
volcanismo canario.
En
Salvaje Pequeña descubrimos, en 1976, un depósito fosilífero cuaternario. Se
trata de una “playa levantada” situada a 18 m sobre el nivel del mar, con
fauna muy similar a la actual.
A
la vista de todo esto y teniendo en cuenta la geomorfología terrestre y
submarina, podemos decir que las Salvajes
actuales son los restos de unas islas mucho mayores (hace 18.000 años tenían
unos 100 km2 de extensión) que están condenadas a desaparecer bajo
el mar en un futuro geológico próximo.
El
clima de este archipiélago, dada su situación geográfica, no difiere mucho
del de Canarias y, por lo tanto, lo podemos catalogar como subtropical marítimo.
Pero es la escasa altura de estas islas (154 m) la que marca las características
climatológicas reales, permitiendo que los vientos dominantes, los alisios
cargados de humedad, las sobrepasen sin condensarse. Esto es lo que ocurre en
Lanzarote y Fuerteventura, en donde escasean las precipitaciones por dicho
motivo.
La
fauna terrestre de Salvajes es pobre, como es obvio, marcada por la pequeña
extensión de las islas y su alejamiento del continente. Entre los vertebrados
destacan las aves marinas, siendo las pardelas (Calonectris
diomedea) las más abundantes y las que han sido objeto de una caza
exhaustiva desde hace siglos para la obtención de aceite, fundamentalmente en
Salvaje Grande. Otra especie muy común, en este caso en Salvaje Pequeña, es el
paiño (Pelagodroma marina), pequeña
ave marina de costumbres nocturnas, que hace sus nidos en madrigueras bajo la
arena.
El
mayor interés, sin embargo, está en los invertebrados terrestres,
especialmente insectos, en los que se da un alto porcentaje de endemismos.
De
la fauna marina destacamos dos moluscos gasterópodos: la lapa de gran tamaño (Patella
candei), muy abundante también en Canarias durante el Cuaternario (ahora es
muy escasa, y se encuentra solamente en las islas orientales) y el “burgado”
endémico (Osilinus selvagensis)
descrito por nosotros.
La
flora de este pequeño archipiélago, al igual que la fauna, presenta gran
afinidad con Canarias y Madeira, aunque, como es lógico, el número de especies
es reducido, dada su pequeña superficie. En Salvaje Grande la flora autóctona
se encuentra refugiada en los acantilados, ya que esta isla ha sufrido la
presencia temporal de algunos colectivos humanos, principalmente canarios, que
pasaban allí largas temporadas en los siglos pasados e introdujeron especies
devastadoras de la flora, como el conejo y la cabra. Por el contrario, Salvaje
Pequeña y La Salvajita, mucho menos antropizadas, albergan interesantes
endemismos como Argyranthemum thalassophilum y la sorprendente tabaiba de La
Salvajita (Euphorbia anacoreta).
El
archipiélago canario, integrado por 7 islas grandes, 4 isletas y varios islotes
y roques, ocupa una superficie oceánica de 7.540 km2, y por lo tanto
es, con mucho, el más extenso de la Macaronesia. También es el que posee la máxima
altura (3.718 m en el Teide, Tenerife), factor que le otorga mayores
posibilidades a la hora de tasar la biodiversidad. Asimismo, es el más próximo
al continente africano (96 km), lo que aumenta la facilidad de poblamiento y
afinidades con este continente.
Sobre
el origen de Canarias se ha vertido mucha tinta. Desde las interpretaciones
mitológicas hasta las teorías actuales del punto caliente o de la fractura
propagante, las controversias han mediado, sobre todo, entre los
“continentalistas” que propugnan que al menos las islas orientales,
Fuerteventura y Lanzarote, estuvieron en otros tiempos unidas al continente
africano, y los “oceanistas” que sostienen el origen oceánico de las islas,
o lo que es lo mismo, que se originaron tras múltiples erupciones submarinas
hasta aflorar definitivamente. Está comprobado que las protoislas canarias
emergieron hace más de 20 millones de años y hoy es posible observar esos
antiguos complejos basales de volcanismo submarino en Fuerteventura, La Gomera,
La Palma y, probablemente, en Tenerife y Gran Canaria. También debemos decir
que en Fuerteventura, y formando parte de ese antiguo complejo basal de origen
submarino, hoy aflorado, se encuentran rocas sedimentarias del Mesozoico (más
de 130 m.a.) de estratificación fina y muy plegada, que contienen fósiles de
ammonites. Algo parecido sucede en la isla de Maio, en Cabo Verde, pero esto no
quiere decir que debamos retrotraer la antigüedad de dichas islas hasta el
Mesozoico, sino que esas rocas son retazos de materiales sedimentarios de la
corteza oceánica subyacente mucho más antiguos, que fueron englobados y
transportados a la superficie en la fase intrusiva y posterior desmantelamiento
de todo el antiguo conjunto insular.

El
volcanismo subaéreo comienza en Canarias hace unos 20 millones de años en
Fuerteventura y Lanzarote, con períodos de máxima actividad entre 14 y 12 m.a.
y entre 5 y 3 m.a., y también largas épocas de calma magmática en las que
operaba la acción erosiva. En cualquier caso, las dataciones radiométricas nos
muestran una progresión de edades desde la isla más occidental (El Hierro, 1
m.a.) hasta las orientales (Fuerteventura, 21 m.a.), que estarían acordes con
el desplazamiento de la placa africana sobre un punto caliente sobre el que está
situado ahora El Hierro. No obstante, se han puesto muchos reparos a esta teoría,
entre ellos las prolongadas interrupciones del volcanismo en Canarias o las últimas
erupciones de Lanzarote. Sin embargo, este archipiélago está atravesando en la
época actual por un período de gran actividad, como lo demuestran las
numerosas erupciones históricas acaecidas en algunas islas, fundamentalmente en
La Palma, Tenerife y Lanzarote. La última ocurrió en 1971 (Teneguía, La
Palma). Con toda probabilidad, en los últimos tiempos también ha habido
actividad volcánica submarina en las proximidades de las islas, como puede ser
el caso del “Volcán del Medio”, monte submarino de 500 m de altura recién
descubierto entre Tenerife y Gran Canaria, que pudiera tener relación con la
falla existente entre ambas islas y con el terremoto de intensidad 5 en la
escala de Richter, ocurrido en 1989, cuyo epicentro se localizó por esa zona.
Un
hecho que quisiéramos resaltar es la gran repercusión paleobiogeográfica que
han tenido las últimas crisis climáticas por las que ha atravesado nuestro
planeta en el poblamiento y evolución de la fauna y flora insular. Si nos
centramos en la época del máximo de la última glaciación cuaternaria (hace
18.000 años) en la que el nivel del mar estaba unos 120 m por debajo del
actual, el panorama geográfico de Canarias era bastante diferente al actual. En
esa época, Fuerteventura, Lanzarote y las isletas e islotes formaban una única
isla, que llamamos Mahan, de unos 5.000 km2 de superficie, 200 km de
largo y a tan sólo 60 km de distancia del continente. Además, entre la península
de Jandía (Fuerteventura) y Gran Canaria existía, también en esa época, la
isla Amanay, de unos 100 km2 de extensión, que en la actualidad es
un banco submarino con abundante pesca, a tan sólo 25 m de la superficie del
mar. Si a todo esto añadimos el sensible incremento en la extensión y altura,
la variación del contorno de las islas en esa época y la consiguiente reducción
de las distancias entre las mismas -al quedar emergidas las plataformas
insulares- nos encontramos con un marco geográfico bien diferente al actual
que, a todas luces, hay que tener en cuenta a la hora de hacer cualquier
planteamiento sobre biogeografía, poblamiento, paleoecología y evolución
insular.
El
clima de Canarias, como el de los restantes archipiélagos, viene determinado
por los vientos dominantes (alisios), las corrientes oceánicas (corriente de
Canarias) y su latitud geográfica. La combinación de estos principales
factores hacen que el clima de este archipiélago se considere entre los más
privilegiados del mundo, con temperaturas medias suaves (20-22 ºC) y con poca
variación anual. Quizás sea el agua el handicap fundamental con el que tienen
que contar las islas, sobre todo Lanzarote y Fuerteventura, en donde las
precipitaciones anuales son escasas (100-150 mm). En general, podemos decir que
la época seca en Canarias transcurre entre los meses de mayo y octubre, que es
cuando comienzan las lluvias, para alcanzar su máximo en noviembre y, a veces,
en primavera.
Un
incidente climático que no hay que olvidar es la presencia en estas islas, con
cierta frecuencia, de vientos procedentes del Sáhara (Harmattan) con polvo en
suspensión, fenómeno conocido en Canarias como “tiempo sur”, que reducen
sensiblemente la visibilidad (calima).
Para
no extendernos demasiado al tratar las variadas e interesantes flora y fauna de
Canarias, trataremos de sintetizar y resaltar los datos más importantes y
significativos. En Canarias hay unas 2000 especies de plantas vasculares (autóctonas
e introducidas asilvestradas), de las que unas 600 son endémicas (casi un
tercio). Son también numerosos en estas islas los endemismos macaronésicos y
las especies comunes a dos o más archipiélagos.
En
el piso basal (0-300 m) se desarrolla el matorral xerófilo caracterizado por
diferentes especies de Euphorbia, las
tabaibas y los cardones, así como otros arbustos, muchos de ellos endémicos
del archipiélago (balos, verodes, magarzas, cardoncillos, etc.). En cotas un
poco más elevadas viven, aunque no con el esplendor de antaño, pequeñas
poblaciones de palmeras (Phoenix
canariensis), sabinas (Juniperus
phoenicea) y en mucha menor cantidad, dragos (Dracaena
draco).
En
el piso montano (400-1800 m) hay que distinguir entre las vertientes norte y sur
de las islas que lo poseen, ya que en la parte baja de la primera, húmeda y
fresca por la incidencia de los alisios, se encuentran los bosques de laurisilva
y fayal-brezal, conocidos en las islas como “monte verde”. La laurisilva
alberga especies arbóreas como el laurel (Laurus
azorica), el palo blanco (Picconia
excelsa) o el viñátigo (Persea
indica), aparte de otras muchas especies arbustivas y herbáceas. Y en la
parte alta (por encima de los 1200 m) se desarrollan los bosques de pino canario
(Pinus canariensis) en medio de un clima mucho más seco. Sin
embargo, en la vertiente sur, más seca y soleada que la norte, solamente está
presente el pinar.
El
piso superior canario o de alta montaña (por encima de los 2.000 m) solamente
lo presentan las islas de Tenerife y La Palma. Está conformado por varias
especies de leguminosas que constituyen el denominado matorral de alta montaña.
Entre ellas destacan la retama del Teide (Spartocytisus
supranubius) en Tenerife y el codeso del pico (Adenocarpus viscosus) en La Palma. También en la alta montaña
canaria crecen especies de alto interés florístico, ya que son pocas las islas
macaronésicas que superan esas alturas, entre ellas el tajinaste rojo (Echium
wildpretti) y la violeta del Teide, que crece por encima de los 2.500 m.
De
la flora fósil de Canarias diremos que los restos vegetales más antiguos (13.5
millones de años) los encontramos hace unos años en el suroeste de Gran
Canaria, en donde pudimos detectar, además de impresiones de hojas de diversas
especies, fragmentos de corteza de pino.
Como
ya hemos comentado, el archipiélago canario es el que presenta mayor
biodiversidad de toda la Macaronesia, consecuencia lógica de sus mayores
dimensiones de superficie y altura y de su proximidad al continente. Por estos
motivos la fauna de Canarias es también, en líneas generales, la más rica.
Dentro de esta gran diversidad destacaremos entre los vertebrados las aves endémicas:
el pinzón azul (Fringilla teydea), la paloma rabiche (Columba junoniae), la paloma turqué (Columba bolli), la tarabilla canaria (Saxicola dacotiae) y el mosquitero (Phylloscopus canariensis). Entre los mamíferos son los murciélagos
y las musarañas los más destacables. De las seis especies de los primeros, que
escasean cada vez más al irse destruyendo su hábitat, dos son compartidas con
Cabo Verde y otras dos con Madeira. Las musarañas (Crocidura spp.) son micromamíferos cuyo descubrimiento en estas
islas ha sido relativamente reciente.
En
cuanto a los reptiles, merecen especial mención los lagartos endémicos,
algunos de considerable tamaño (más de 50 cm), como Gallotia
simonyi de la isla de El Hierro, Gallotia
stehlini de Gran Canaria y la especie recién descubierta en Tenerife Gallotia
sp. Otros reptiles que presentan diversas razas o subespecies en estas islas
son las lisas (Chalcides spp.) y los
gecónidos (Tarentola spp.) conocidos
aquí como perenquenes.
No
menos interesante resulta la fauna fósil de vertebrados, fiel testimonio de un
pasado insular sorprendente, como lo demuestra la presencia de aves gigantes en
el Mioceno superior (6 m.a.) de Lanzarote y de grandes tortugas terrestres cuyos
restos han aparecido en esta isla –en el mismo yacimiento de las aves
gigantes- en el Plioceno de Fuerteventura (Geochelone
sp.) y en el Pleistoceno de Gran Canaria (G.
vulcanica) y de Tenerife (G. burchardi).
Los lagartos también han tenido representantes gigantescos en el pasado de
estas islas. Gallotia goliath, que
vivió en Tenerife y La Gomera hasta hace unos pocos miles de años, alcanzaba
1.5 m de longitud total. Pero quizás lo más sorprendente en lo que a reptiles
fósiles se refiere, es el reciente hallazgo, por nuestra parte, de una vértebra
de serpiente (bóido) en el mismo depósito calcarenítico de las aves gigantes
de Lanzarote, que viene a complicar aún más la siempre controvertida explicación
sobre el origen y poblamiento de las Canarias orientales.
El
archipiélago de Cabo Verde, situado a unos 500 km al oeste del Cabo Verde (de
ahí su nombre) próximo a Dakar -la capital de Senegal- y separado de Canarias
1300 km, representa a la Macaronesia árida, escasa en agua. Cabo Verde
comprende diez islas, dos isletas y varios islotes y roques más pequeños, que
se distribuyen desde antiguo en dos grupos: Barlovento y Sotavento. Sin embargo,
resulta más adecuado, según la realidad geográfica y geomorfológica, dividir
a este archipiélago en tres grupos de islas: grupo noroccidental, que comprende
a Santo Antao, San Vicente, Santa Luzía, las isletas Branco y Raso, y San
Nicolau; el grupo oriental, que incluye a Sal, Boa Vista y Maio; y finalmente,
al grupo sur quedarían adscritas Santiago, Fogo, los islotes Rombos y Brava. En
total ocupan una extensión de poco más de 4.000 km2, siendo la
mayor Santiago, con una superficie de 990 km2 y la más pequeña
Santa Luzía con 35.
Como
decíamos, la geomorfología -que viene casi siempre determinada por la antigüedad-
es muy variable de una isla a otra, especialmente entre las del grupo oriental,
bajas y arenosas, y las occidentales, más altas y verdes.
De
origen netamente oceánico, estas islas fueron consideradas como las más
antiguas de la Macaronesia, merced al afloramiento en Maio de rocas
sedimentarias mesozoicas, al igual que sucede en la región del Puerto de la Peña
en Fuerteventura. Estudios recientes han demostrado que estos sedimentos marinos
han sido transportados a la superficie desde la corteza oceánica, de la que
formaban parte en la primera etapa -fuertemente intrusiva- de la formación de
estas islas.
El
volcanismo reciente queda bien patente en la isla de Fogo, en donde la intensa
actividad magmática de los últimos tiempos geológicos han conformado el único
estrato-volcán del Archipiélago, el “Pico do Fogo”, de 2830 m de altura,
que se asienta sobre una espectacular caldera cuyas paredes verticales llegan a
alcanzar los 1000 m de desnivel. La última manifestación de este volcán
ocurrió en 1995, quedándose la lava emitida retenida en el interior de la
Caldera. No sucedió así en la anterior erupción, en 1951, cuyas coladas lávicas
la sobrepasaron por la cara suroeste y discurrieron pendiente abajo.
De
los avatares geotectónicos y erosivos que ha sufrido este archipiélago dan fe
las extensas plataformas insulares submarinas que unen las islas de Maio, Boa
Vista y Sal, al igual que sucede con las del grupo noroccidental, en donde casi
nunca se superan los 50 m de profundidad, llegando –incluso- hasta los 9 m y
suponiendo un peligro para la navegación interinsular, como es el caso del
banco de Joao Valente, entre Maio y Boa Vista.
En
el aspecto paleontológico diremos que las más ricas son las islas orientales,
que a su vez son las más antiguas, junto a Santiago y algunas del grupo
noroccidental. Niveles marinos importantes, a diversas altitudes, y paleodunas
consolidadas que se distribuyen a lo largo de casi toda la costa de estas islas,
constituyen depósitos calcareníticos fosilíferos, muchas veces ricos en fauna
-fundamentalmente moluscos- y flora (algas calcáreas) en parte ya desaparecidas
de las islas.
También
en relación con los movimientos eustáticos del nivel de mar están las
terrazas marinas, que a veces se acercan o sobrepasan los 100 m de altitud, bien
patentes hoy en día en los acantilados y extensas llanuras litorales de las
islas anteriormente mencionadas, fundamentalmente en Boa Vista y Sal, en donde
es posible seguir estos niveles a lo largo de kilómetros. Según ésto podemos
suponer que, dada la escasa altura de estas islas (rara vez superan los 50 m,
excepto algunas montañas o conos volcánicos) en los períodos cálidos del
Cuaternario y Cenozoico, su superficie se vería sensiblemente reducida,
convirtiéndose, probablemente, en varios islotes separados por estrechos brazos
de mar. Resultan obvias, por lo tanto, las implicaciones biogeográficas y de
evolución insular que de ello derivan.
Cabe
destacar también, en lo que se refiere a vertebrados fósiles, la presencia de
restos de tortugas terrestres en el interior de la caldera de Pedra Lume en la
isla de Sal. Actualmente en estudio, estos fósiles pueden aportar nuevos datos
para la interpretación del poblamiento faunístico insular.
El
clima templado de Cabo Verde es una consecuencia de su condición atlántica y
de su latitud (entre 14ºN y 18ºN). A pesar de que la temperatura máxima en
este archipiélago casi nunca sobrepasa los 30ºC, la sensación de calor es muy
superior, debido a su situación tropical. Quizás lo que más condiciona la
climatología y, por ende, la vida de estas islas, es la pluviosidad -que es muy
escasa e irregular- especialmente en las más bajas y orientales. Las estaciones
apenas se dejan sentir y es la época de lluvias, entre julio y octubre -cuando
el frente intertropical se desplaza hacia el norte- el momento en que se aprecia
un cambio visible, al cubrirse la árida superficie del archipiélago de un
manto vegetal que “devuelve la vida” a estas necesitadas islas.
La
flora del archipiélago, como consecuencia de su situación geográfica, muestra
bastante afinidad con la paleotropical y algo menos con la del Mediterráneo y
de los otros archipiélagos de la Macaronesia. A pesar de que la influencia de
la flora oesteafricana es muy patente, los endemismos macaronésicos son allí
frecuentes, como los Aeonium
(verodes), Echium (tajinastes), Limonium
(siemprevivas) o Lotus
(corazoncillos). Tampoco es raro encontrar en estas islas Sonchus (cerraja), Teline (retama),
Launaea (aulaga), Phoenix
(palmera) o Dracaena (drago) aunque este último, antiguamente muy abundante en
algunas islas como San Nicolau, prácticamente ha desaparecido debido a la
sobreexplotación a que se vió sometido para la extracción de su savia (sangre
de drago) en los primeros siglos de la colonización. Algunas especies de la
flora liquénica como Roccella spp.
(orchilla) también fueron objeto de un intenso comercio en esa época.
En
todo caso, lo más resaltable de la flora caboverdiana es su profunda alteración
por la mano del hombre: desforestación, introducción de especies alóctonas,
etc. Por ese motivo ahora el paisaje vegetal está dominado por leguminosas arbóreas
(Prosopis, Acacia, etc.),
piteras (Fourcraea) y otras muchas
plantas introducidas.
Al
igual que sucede con la flora, la fauna de este archipiélago no es, en general,
muy afin a la de las demás islas macaronésicas. No obstante, en lo que a la
avifauna se refiere, se dan bastantes similitudes, aunque, como es lógico, con
aportes tropicales. Por lo tanto, podemos observar en Cabo Verde muchas especies
comunes con Canarias, como es el caso del gorrión (Passer
hispanoliensis), la curruca tomillera (Sylvia
conspicillata), la paloma bravía (Columba
livia) o el capirote (Sylvia
atricapilla). También es frecuente ver allí aves que desgraciadamente ya
han desaparecido de Canarias o están a punto de hacerlo, como el alimoche (Neophron
percnopterus) y el milano real (Milvus
milvus). Pero son los reptiles los que quizás merezcan una especial atención,
ya que de las siete especies presentes en las islas, cuatro son endémicas (60%)
y entre ellas los escíncidos (lisas) -representados por los géneros Mabuya y Macroscincus- son
los más diferenciados. Macroscincus
es un género endémico de Cabo Verde, cuya una única especie, M.
coctei, alcanzaba hasta 60 cm de longitud total y habitaba en Santa Luzía
y, sobre todo, en las isletas Branco y Raso. Desgraciadamente estamos hablando
en pasado, porque ya se da por extinguido este interesantísimo género. Y ésto
ha ocurrido, prácticamente ante nuestros ojos, en las últimas décadas.
Del
resto de la fauna destacaremos también el gran número de insectos endémicos,
sobre todo en algunos grupos, pero, en general, siempre mostrando una clara
influencia africana, mayor que en los otros archipiélagos macaronésicos.
Con
respecto a la fauna marina cabe resaltar la presencia de fondos coralinos que,
si bien no llegan a constituir verdaderos arrecifes, sí conforman ecosistemas
particulares, con la incorparación de elementos tropicales que no están
presentes en el resto de la Macaronesia. Sin embargo, son muchas las especies
que podemos considerar como panmacaronésicas.
ENCLAVE CONTINENTAL
Desde
el punto de vista biogeográfico existe una relación natural entre los archipiélagos
macaronésicos -fundamentalmente las islas Canarias orientales y Cabo Verde- y
una franja costera atlántica de miles de kilómetros, que va desde el suroeste
de la Península Ibérica, pasando por el sur de Marruecos, Sáhara Occidental y
Mauritania, hasta el norte de Senegal.
La
relación de Lanzarote y Fuerteventura está más que explicada por la corta
distancia al continente africano (96 km), que fue aún más corta durante las épocas
glaciales del Cuaternario (hace 18.000 años era de 60 km). Las afinidades son
apreciables especialmente en el paisaje vegetal, muy parecido al de los pisos
basales de las islas orientales. En esas zonas, un elevado número de especies
vegetales y algunas animales (insectos) son comunes -y exclusivas- de ambas
regiones, y bastantes más que son vicariantes. Es por esto por lo que se habla
de enclave continental macaronésico. Y dentro de esta extensísima franja
costera africana, existe un espacio
reducido pero muy representativo, el Cabo Guir, a unos 30 km al norte de Agadir
(Marruecos), que es un ejemplo magnífico de relaciones biogeográficas entre
regiones próximas. Allí podemos ver tabaibas y cardones (Euphorbia
spp.), aulagas (Launaea),
cornicales (Periploca), orobales (Withania),
verodes (Senecio), bejeques (Aeonium),
tasaigos (Rubia) y cerrajas (Sonchus),
entre otros, formando parte de una misma comunidad vegetal.
El
clima de esta franja continental es árido, como es natural, y parecido al de
las zonas costeras de las Canarias orientales. También allí la influencia oceánica
(corriente fría de Canarias) se deja sentir y suaviza, de alguna manera, las
temperaturas. De igual manera los vientos alisios, frescos y húmedos, influyen
y propician la humedad y las escasas precipitaciones necesarias para la
supervivencia de estas comunidades biológicas, pese a la proximidad del
desierto. Estos vientos soplan paralelos a la costa y apenas se introducen en el
interior, debido a la barrera que representa la cordillera del Atlas, y es
precisamente en esas montañas del Sus marroquí, en la frontera del desierto,
donde recientemente se descubrió para la ciencia una numerosa población de
dragos (Dracaena) junto a otras
formaciones vegetales arbóreas que también viven, de manera relíctica, en
algunas de las islas macaronésicas. Es el caso de los bosques de sabina,
acebuche, lentisco, etc.
Pero
son las tabaibas y cardones (Euphorbia
spp.) las que podemos considerar como “marcadoras” a la hora de hacer
estudios sobre la distribución geográfica o de la extensión del enclave
macaronésico continental, ya que son más visibles, y fácilmente detectables
en el campo, que otras plantas de menor porte que conviven con ellas.
De
la fauna de este enclave no podemos decir lo mismo que de la flora. Son muy
pocas las especies comunes con los archipiélagos. Pensamos que esto es debido,
entre otras causas, a la mayor movilidad y amplia valencia ecológica que
presentan los animales con respecto a las plantas. En este sentido, es Cabo
Verde el archipiélago que alberga mayor número de especies afines, sobre todo
insectos, a pesar de estar mucho más alejado del continente que Canarias. La
explicación puede estar en la latitud tropical que ocupan estas islas, que hace
que su clima se corresponda más con la región costera continental que tienen
enfrente. Este hecho es extensible también a la fauna marina, que presenta un
alto porcentaje de especies tropicales comunes, aunque también abundan los
endemismos y las especies panmacaronésicas.
Abdel-Monem,
A.A., Fernández, L.A. y Boone, G.M., 1975. K-Ar
ages from the eastern Azores group (Santa María, Sâo Miguel and the
Anguita, F. y Hernán, F., 1975. A
propagating fracture model versus a hot spot origin for the
Báez, M. y Sánchez-Pinto, L., 1983. Islas de fuego y agua.
Canarias, Azores, Madeira, Salvajes, Cabo Verde. Edirca, 184 pp.
Barahona,
F., Rage, J.C. y García-Talavera, F., 1998.
The first record of snakes on the
Blondel,
J., 1997. Les
adaptations du vivant, in: Îles. Vivre
entre ciel el mer. Muséum
National D´Histoire Naturelle.
Boekschoten, G.J. y Borel Best, M., 1988.
Fossil and recent shallow water corals from the
Bravo,
T., 1983. El origen de las
islas Canarias, in: Canarias, origen y
poblamiento. pp. 15-24. Círculo de Estudios Sociales de Canarias. Queimada
Ed. Madrid.
Carracedo,
J.C. et.al., 1998. Origen y
evolución del volcanismo de las islas Canarias, in: Ciencia
y Cultura en Canarias, pp. 67-89. Museo de la Ciencia y el Cosmos, O.A.M.C.,
Cabildo de Tenerife.
Francisco
Ortega, J., Jansen, R.K., Crawford, D.J. y Santos, A., 1995. Chloroplast
DNA evidence for intergeneric relationships of the Macaronesian endemic genus Argyranthemum
(Asteraceae). Sistematic
Botany 20: 413-422.
Francisco
Ortega, J., Jansen, R.K. y Santos Guerra, A., 1996. Chloroplast
DNA evidence of colonization adaptative radiation, and hybridization in the
evolution of the Macaronesian flora. Proceedings
of the National
Galopim
de Carvalho, A.M. y Brandao, J.M., 1991. Geología do arquipélago
da Madeira. Mus. Nac.
De Historia Natural. Universidade de Lisboa, 171 pp., 32 lam.
García-Talavera,
F., 1978. Sobre el
Cuaternario marino de la isla Selvagem Pequena, in: Contribución al estudio de la Historia Natural de las islas Salvajes.,
pp. 37-44. Museo de Ciencias Naturales, Aula de Cultura del Cabildo de Tenerife.
García-Talavera,
F., 1983. Los
moluscos gasterópodos anfiatlánticos. Estudio paleo y biogeográfico de
las especies bentónicas litorales. Publ. Univ. La Laguna. Monografías, nº 10.
352 pp. 7 lam.
García-Talavera,
F., 1990. Aves gigantes en
el Mioceno de Famara (Lanzarote). Rev.
Acad. Canar. Cienc., II, 71-79.
García-Talavera,
F., 1997. Las Canarias
orientales y vecina costa africana en el Holoceno. ERES (Arqueología), Vol. 7 (1): 55-63.
García-Talavera,
F., Sánchez-Pinto, L.y Socorro S., 1995. Vegetales
fósiles en el complejo traquítico-sienítico de Gran Canaria. Rev.
Acad. Canar. Cienc., VII (2, 3 y 4), 77-91.
Granjon,
L. y Barrau, J., 1997. Comment coloniser les îles ?, in: Îles.
Vivre entre ciel et mer. Muséum National D´Histoire Naturelle.
Grant, P.R., 1998. Evolution on
Hunter, P.M., Searle, R.C. y Laughton, A.S., 1983. Bathymetry
of the N.E. Atlantic, Sheet 5: Continental margin off N.W. Africa. Institute
of Oceanographic Sciences,
Hutterer, R., García-Talavera, F., López-Martínez, N. y Michaux, J.,
1998. New
chelonian eggs from the Tertiary of Lanzarote and Fuerteventura, and a review of
fossil tortoises of the
La
Roche, F. y Rodríguez-Piñero, J.C., 1994.
Aproximación al número de taxones de la flora vascular silvestre de los
archipiélagos macaronésicos. Rev. Acad.
Canar. Cienc., VI (2,3,4): 77-98.
Machado,
A., 1998. Biodiversidad.
Un paseo por el concepto y las islas Canarias. Cabildo
de Tenerife. 67 pp.
Meco, J., 1988. The emergent littoral deposits in
Menard, H.W., 1986. Islands.
Scientific American Library.
Mitchell-Thomé, R.C., 1983. The
Mitchell-Thomé, R.C., 1985. Radiometric studies in Macaronesia. Bol.
Mus. Mun. Funchal, 37
(167): 52-85.
Nicolás
de, J.P., Fernández-Palacios, J.M., Ferrer, F.J. y Nieto, E., 1989. Inter-island
floristic similarities in the Macaronesian region. Vegetation 84: 117-125.
Nunn, P.D., 1994. Oceanic
Petit-Maire,
N., Delibrias, G., Meco, J., Pomel, S. y Rosso, J.C., 1987.
Paléoclimatologie des Canaries Orientales (Fuerteventura). C.R.
Acad. Sc. Paris.
303,
Sér. II, 13: 1241-1246.
Sergel, R. y Báez, M., 1990. On the Biotic Diversity of Eastern Atlantic
Islands and its implication for the Theory of Island Biogeography. Courier Forsch.-Inst. Senckenberg, 129: 25-41.
Serralheiro,
A., 1970. A
geologia da Ilha de Maio
(Cabo Verde). Junta de Investigaçoes do Ultramar, Lisboa., 103 pp.
Schmincke, H.U., 1973. Magmatic evolution and tectonic regime in the Canary,
Madeira and
Schmincke, H.U., 1976. The geology of the
Stillman, C.J., et. al., 1982. The geological history of
Vigne, J.D., 1997. L`emprise
de l`homme depuis le préhistoire, in: Îles.
Vivre entre ciel et mer. Muséum National D´Histoire Naturelle. Paris.